 Add to Chrome
Add to Chrome Add to Firefox
Add to Firefox Add to Edge
Add to EdgeLocal search for valued constraint satisfaction parameterized by treedepth
May 20, 2024Sometimes local search algorithms cannot efficiently find even local peaks. To understand why, I look at the structure of ascents in fitness landscapes from valued constraint satisfaction problems (VCSPs). Given a VCSP with a constraint graph of treedepth $d$, I prove that from any initial assignment there always exists an ascent of length $2^{d + 1} \cdot n$ to a local peak. This means that short ascents always exist in fitness landscapes from constraint graphs of logarithmic treedepth, and thus also for all VCSPs of bounded treewidth. But this does not mean that local search algorithms will always find and follow such short ascents in sparse VCSPs. I show that with loglog treedepth, superpolynomial ascents exist; and for polylog treedepth, there are initial assignments from which all ascents are superpolynomial. Together, these results suggest that the study of sparse VCSPs can help us better understand the barriers to efficient local search.
Nothing makes sense in deep learning, except in the light of evolution
May 20, 2022


Deep Learning (DL) is a surprisingly successful branch of machine learning. The success of DL is usually explained by focusing analysis on a particular recent algorithm and its traits. Instead, we propose that an explanation of the success of DL must look at the population of all algorithms in the field and how they have evolved over time. We argue that cultural evolution is a useful framework to explain the success of DL. In analogy to biology, we use `development' to mean the process converting the pseudocode or text description of an algorithm into a fully trained model. This includes writing the programming code, compiling and running the program, and training the model. If all parts of the process don't align well then the resultant model will be useless (if the code runs at all!). This is a constraint. A core component of evolutionary developmental biology is the concept of deconstraints -- these are modification to the developmental process that avoid complete failure by automatically accommodating changes in other components. We suggest that many important innovations in DL, from neural networks themselves to hyperparameter optimization and AutoGrad, can be seen as developmental deconstraints. These deconstraints can be very helpful to both the particular algorithm in how it handles challenges in implementation and the overall field of DL in how easy it is for new ideas to be generated. We highlight how our perspective can both advance DL and lead to new insights for evolutionary biology.
Steepest ascent can be exponential in bounded treewidth problems
Dec 02, 2019We investigate the complexity of local search based on steepest ascent. We show that even when all variables have domains of size two and the underlying constraint graph of variable interactions has bounded treewidth (in our construction, treewidth 7), there are fitness landscapes for which an exponential number of steps may be required to reach a local optimum. This is an improvement on prior recursive constructions of long steepest ascents, which we prove to need constraint graphs of unbounded treewidth.
Complexity of evolutionary equilibria in static fitness landscapes
Aug 23, 2013

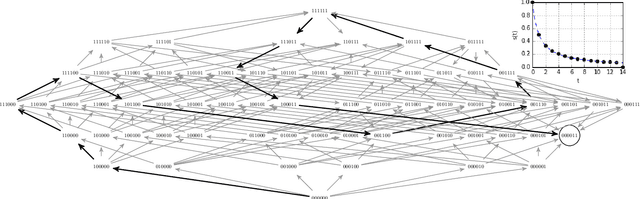

A fitness landscape is a genetic space -- with two genotypes adjacent if they differ in a single locus -- and a fitness function. Evolutionary dynamics produce a flow on this landscape from lower fitness to higher; reaching equilibrium only if a local fitness peak is found. I use computational complexity to question the common assumption that evolution on static fitness landscapes can quickly reach a local fitness peak. I do this by showing that the popular NK model of rugged fitness landscapes is PLS-complete for K >= 2; the reduction from Weighted 2SAT is a bijection on adaptive walks, so there are NK fitness landscapes where every adaptive path from some vertices is of exponential length. Alternatively -- under the standard complexity theoretic assumption that there are problems in PLS not solvable in polynomial time -- this means that there are no evolutionary dynamics (known, or to be discovered, and not necessarily following adaptive paths) that can converge to a local fitness peak on all NK landscapes with K = 2. Applying results from the analysis of simplex algorithms, I show that there exist single-peaked landscapes with no reciprocal sign epistasis where the expected length of an adaptive path following strong selection weak mutation dynamics is $e^{O(n^{1/3})}$ even though an adaptive path to the optimum of length less than n is available from every vertex. The technical results are written to be accessible to mathematical biologists without a computer science background, and the biological literature is summarized for the convenience of non-biologists with the aim to open a constructive dialogue between the two disciplines.